




 下载:
下载:




-
森林是陆地生态系统的主体,其生物量和净生产力占整个陆地生态系统的86%和70%,对全球碳平衡起着十分重要的作用[1-3]。生物量作为植物的基本生物学特征和功能性状之一,是物质和能量积累的基本体现[4-5]。掌握森林中各类树木生物量的分配特征有助于了解碳储量和碳平衡的动态变化。测定树木生物量的方法通常有皆伐法、平均标准木法、径级标准木法和异速生长模型法等[6-8]。直接测定法获取生物量的方法精度虽然较高,但是费时、费力,且对生态环境破坏较严重[9-10];生物量异速生长模型将简单、易获取的变量与树木生物量结合,为森林生态系统生物量和净生产力的估测提供了一种普遍且可靠的方法[6,11-13]。由2014年森林资源二类调查数据统计可知:天山雪岭云杉Picea schrenkiana约占新疆山地森林总面积的59.0%,是新疆山地森林中分布最广、蓄积量最大的森林生态树种,对新疆山地水源涵养、水土保持,以及林区生态系统的碳平衡发挥着不可代替的作用。近年来,张绘芳等[14]从年龄、径阶、林分郁闭度等角度,分析了西伯利亚云杉Picea obovata各组分器官生物量比例变化规律;白志强等[15]利用阿尔泰山西伯利亚云杉各器官生物量实测数据,分别构建了各器官(干、枝、叶、根、地上及整株生物量)的异速生长模型,均取得了较为理想的效果。本研究通过整株收获法调查了30株天山雪岭云杉各器官的分配规律,并基于胸径、树高因子建立各器官的生物量异速生长模型,旨在为生物量估测及异速生长模型优化提供理论依据,为雪岭云杉生物量及碳储量估算提供有效的研究方法。
-
天山雪岭云杉主要分布在新疆天山海拔1 600~2 800 m的中山地带,属温凉半湿润、半干旱针叶林气候区。山区年均气温为0.7 ℃,年降水量为 200~1 000 mm,多集中在6−8月,年蒸发量980~1 150 mm,年均相对湿度65%。本研究选择地处新疆天山山脉中部、西部、东部的3个林区作为研究区,其中板房沟林场(43°24′48.3″~43°26′17.9″N,87°27′28.5″~87°28′47.7″E)位于乌鲁木齐县,地处天山山脉中段北坡,海拔1 908~2 960 m;哈密林场(42°47′10″~43°56′48″N,91°22′43″~94°49′44″E)位于哈密市,地处天山山脉东端;昭苏林场(42°26′13″~43°02′42″N,80°17′15″~81°42′21″E)位于伊犁哈萨克自治州昭苏县,地处天山山脉西段。
-
2013年7−8月从各林场雪岭云杉的分布下限至分布上限每隔200 m分别选取自然生长状况良好、具有代表性的林分布设样地,每个林场布置5块20 m × 20 m临时样地。测量样地内土壤厚度、坡位、坡向、坡度、海拔、下木盖度、下木高度等7个立地因子及平均年龄、平均树高、平均胸径、郁闭度、疏密度、每公顷蓄积量等6个测树因子;根据各样地每木检尺的结果计算样木平均胸径和平均树高,确定2株最接近的雪岭云杉作为标准木,并对30株标准木整株挖掘。
采用分层分割法测定树干生物量,按2 m区分段,称量各区分段的树干鲜质量[16];将树冠分为3 层,每层选取3~5个标准枝,分别称枝、叶的鲜质量;各样品带回室内采用烘干法测定其含水量。采用全挖法分别测定树根(粗根>5 cm、大根2~5 cm、小根≤2 cm)的鲜质量,并测各样品含水量。所有样品带回室内,85 ℃ 烘干至恒量后,推算标准木各器官干质量(kg),汇总得到枝、叶、干和根生物量。其中,地上生物量=枝生物量+叶生物量+干生物量;总生物量=地上生物量+根生物量。
-
相对生长模型对单株树木生物量的拟合和估计精度均有很大的优势[17-19],胸径及树高是计算单株树木生物量的重要变量,在野外调查中很容易获取。本研究主要基于胸径和树高因子,利用Matlab中的拟合工具箱对单株雪岭云杉的各器官生物量进行拟合。
$${W=a{D^b}}\text{;} $$ (1) $$ {W = a{H^b}}\text{;} $$ (2) $${W = a{{({D^2}H)}^b}}\text{;} $$ (3) $${W = a{D^b}{H^c}}\text{;} $$ (4) $$ W = a{({D^3}/H)^b}\text{。} $$ (5) 式(1)~式(5)中:W为
各器官生物量(kg),D为胸径(cm),H为树高(m),A为年龄(a),a、b、c为拟合系数。利用调整决定系数 $R_{\rm{adj}}^2$ 和平均预测误差EMP(%)估计模型拟合优度。公式如下:$$R_{\rm{adj}}^2 = 1 - \frac{{\sum\limits_{i = 1}^n {{{({y_i} - \mathop {{y_i}}\limits^ \wedge )}^2}} }}{{\sum\limits_{i = 1}^n {{{({y_i} - \mathop {{y_i}}\limits^ - )}^2}} }}\text{;}$$ (6) $$E_{\rm{MP}} = \frac{{100}}{n}\sum\limits_{i = 1}^n {\frac{{\left| {{y_i} - \mathop {{y_i}}\limits^ \wedge } \right|}}{{{y_i}}}}\text{。} $$ (7) 式(6)~式(7)中:
${y_i}$ 、$\mathop {{y_i}}\limits^ \wedge $ 分别为各器官生物量的实测值和预测值,$\mathop {{y_i}}\limits^ - $ 为各器官生物量的平均值。 -
作为森林经营、组织木材生产的最小单位和调查设计的基本单位,研究中通常将立地条件、林分因子、采伐方式、经营措施相同和集材系统一致的林分划为1个小班。本研究利用板房沟林场、哈密林场和昭苏林场2014年森林资源二类调查数据,以起源(天然林)和优势树种(雪岭云杉)为筛选条件,3个林场分别划出1 162、873个和2 756个小班;获得所有4 791个小班的土壤厚度、坡位、坡向、坡度、海拔、下木盖度、下木高度等7个立地因子及平均年龄、平均树高、平均胸径、郁闭度、疏密度、每公顷蓄积量等6个测树因子。利用上述单株生物量拟合公式及筛选出的各样地平均树高和平均胸径,计算出小班水平上的单株雪岭云杉平均生物量。利用随机森林法确定13个因子对整株生物量的相对重要性,并排序。其中:坡向、坡位和土壤厚度3个定性指标根据国家森林资源连续清查技术规定对其进行量化[20](表1)。
表 1定性指标量化标准
Table 1.Qualitative indicator quantitative standard
评价标准 量化标准 评价标准 量化标准 坡向 坡位 土壤厚度/cm 坡向 坡位 土壤厚度/cm 无坡向 − − 9 东南 下 − 4 西北 − − 8 东 中 <30(薄) 3 西 − − 7 东北 上 30~59(中) 2 西南 平地 − 6 北 脊 ≥60(厚) 1 南 谷 − 5 说明:“−”表示无相关定性等级 -
本研究所选的30株雪岭云杉单株总生物量实测值为12.04~2 014.34 kg·株−1,地上生物量和地下生物量大分别为10.16~1 475.17 和1.88~539.18 kg·株−1,根冠比为0.08~0.55。随胸径增加,雪岭云杉整株生物量、地上生物量、地下生物量均呈增加趋势,但根冠比变化不明显(表2)。地上生物量与地下生物量的Person相关系数为0.918 2(P<0.001),即两者存在极显著强正相关关系。由图1可知,两者为幂函数关系,拟合方程为y=0.263 7x1.026 7。对地上生物量和地下生物量进行以10为底的对数转换,并进行回归分析和配对样本t检验,结果发现:雪岭云杉地上生物量与地下生物量显著相关且存在极显著差异(P<0.01),拟合方程y=1.027 6x−0.579,说明雪岭云杉地上生物量与地下生物量存在显著异速生长关系(P<0.01)。

图 1雪岭云杉地上与地下生物量的关系
Figure 1.Relationship between aboveground and underground biomass ofP. schrenkiana
表 2天山雪岭云杉单木各器官生物量特征表
Table 2.Characteristic table of individual organ biomass ofP. schrenkianain Tianshan Mountains
径级/cm 整株生物量/(kg·株−1) 地上生物量/(kg·株−1) 地下生物量/(kg·株−1) 根冠比 区间 平均 区间 平均 区间 平均 区间 平均 6.5~20.0 12.04~183.49 70.29 10.16~169.45 55.90 1.88~36.14 14.39 0.08~0.51 0.31 21.0~38.0 124.43~541.62 338.03 99.78~400.49 254.29 24.65~141.12 83.74 0.21~0.54 0.33 41.0~60.0 552.61~1 395.75 918.82 441.28~1 101.87 687.17 111.33~376.77 231.65 0.18~0.55 0.34 61.0~81.0 1 830.42~2 014.34 1 922.38 1 455.44~1 475.17 1 465.31 374.98~539.18 457.08 0.26~0.37 0.31 从整株水平来看(图2):雪岭云杉干、根、枝和叶各器官的生物量相对分配占比分别为(58.86±7.77)%、(24.15±6.37)%、(13.03±4.80)%、(5.96±2.65)%,其生物量分配均表现为干>根>枝>叶;t检验发现:雪岭云杉各器官的生物量分布存在显著性差异(P<0.01)。由图3可知:雪岭云杉干、枝、叶、根各器官基本符合生物量随其胸径增长而增大的趋势。
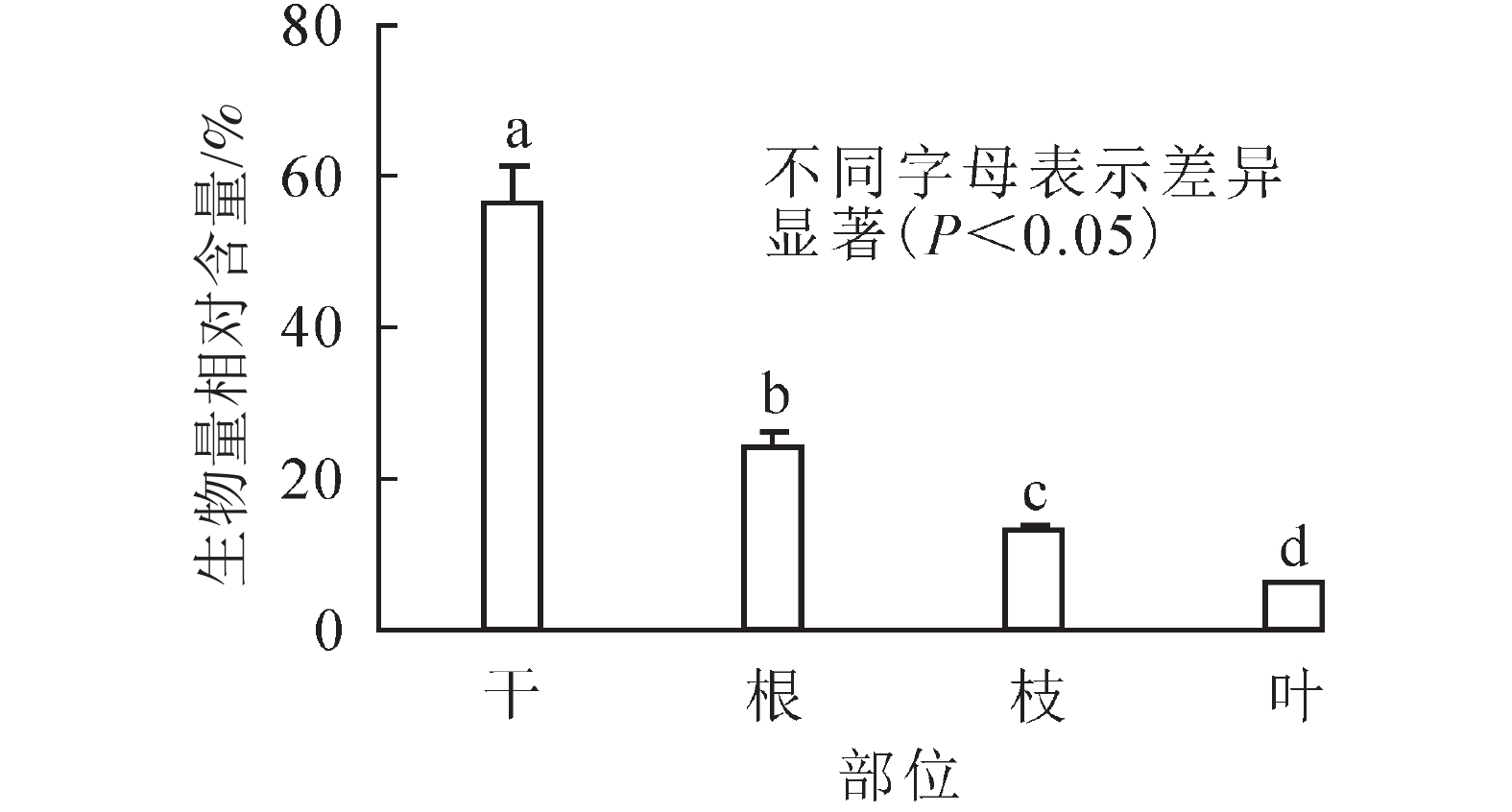
图 2雪岭云杉不同器官生物量相对分配比例
Figure 2.Relative allocation ratio of biomass in different organs ofP. schrenkiana
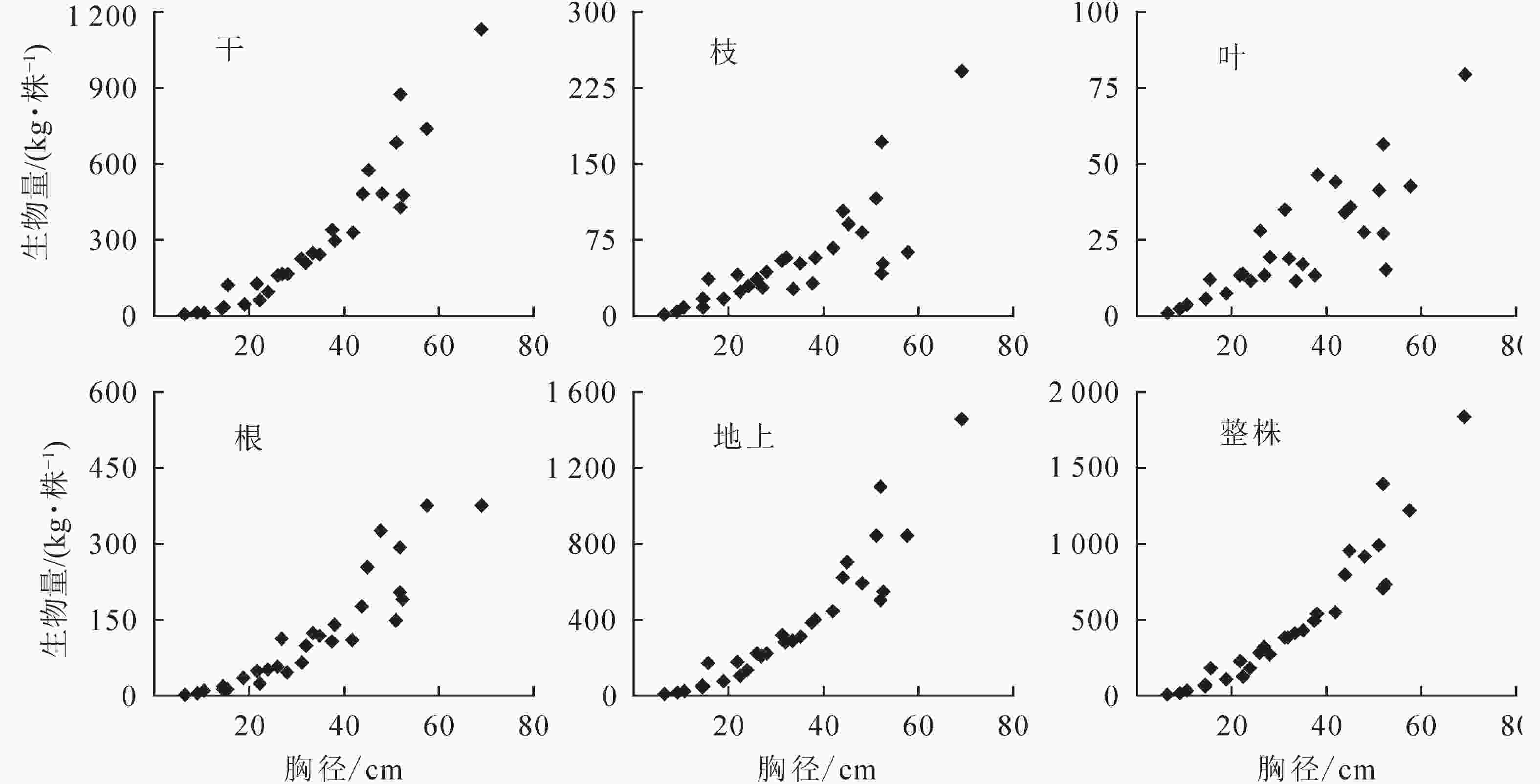
图 3天山雪岭云杉单木各器官生物量随径级变化图
Figure 3.Variation of individual organ biomass with diameter ofP. schrenkianain Tianshan Mountains
-
对天山雪岭云杉的各器官(干、枝、叶和根)、地上生物量及整株生物量进行非线性生物量估测,依据最大相关系数(R2)以及最小平均预测误差(EMP)判断模型优劣(表3)。其中,树干生物量最优模型为W=0.088 5D0.625H1.938,叶生物量最优模型为W=0.035 8D0.229H1.881,根生物量最优模型为W=0.100 6(D2H)0.697,地上生物量最优模型为W=0.104 8D0.551H2.042,整株生物量最优模型为W=0.184 3D0.758H1.708。对树枝生物量模型的拟合发现,无最大R2及最小平均预测误差的组合,故选择最大R2为最优模型判断指标,即树枝的最优生物量模型为W=0.004 9D0.252H2.736。
表 3新疆天山雪岭云杉各器官生物量5种非线性生物量估测模型
Table 3.Five nonlinear biomass estimation models ofP. schrenkianaorgans in Tianshan forest areas
器官 生物量模型 拟合公式 R2 EMP/% 树干 W=aDb W=0.476 6D1.803 0.926 6 38.98 W=aHb W=0.050 6H2.859 0.951 9 25.19 W=a(D2H)b W=0.251 7(D2H)0.695 0.947 5 30.69 W=aDbHc W=0.088 5D0.625H1.938 0.963 1 23.53 W=a(D3/H)b W=1.389 0(D3/H)0.706 0.876 7 63.69 树枝 W=aDb W=0.073 6D1.835 0.768 0 34.49 W=aHb W=0.003 8H3.114 0.830 7 43.52 W=a(D2H)b W=0.033 8(D2H)0.719 0.796 0 35.15 W=aDbHc W=0.004 9D0.252H2.736 0.832 5 42.78 W=a(D3/H)b W=0.139 0(D3/H)0.769 0.709 9 36.41 树叶 W=aDb W=0.167 2D1.395 0.753 6 40.68 W=aHb W=0.030 5H2.205 0.800 9 31.27 W=a(D2H)b W=0.100 9(D2H)0.539 0.777 1 35.39 W=aDbHc W=0.035 8D0.229H1.881 0.802 8 30.69 W=a(D3/H)b W=0.281 5(D3/H)0.581 0.687 9 47.56 树根 W=aDb W=0.178 8D1.826 0.904 4 37.43 W=aHb W=0.032 3H2.722 0.857 8 36.64 W=a(D2H)b W=0.100 6(D2H)0.697 0.908 7 31.85 W=aDbHc W=0.101 9D1.405H0.681 0.908 7 31.92 W=a(D3/H)b W=0.422 7(D3/H)0.741 0.880 0 53.82 地上生物量 W=aDb W=0.635 8D1.786 0.915 7 29.52 W=aHb W=0.064 1H2.854 0.948 4 24.17 W=a(D2H)b W=0.332 3(D2H)0.69 0.938 3 22.83 W=aDbHc W=0.104 8D0.551H2.042 0.957 3 19.09 W=a(D3/H)b W=1.701 0(D3/H)0.708 0.860 3 46.40 总生物量 W=aDb W=0.813 0D1.796 0.938 8 27.08 W=aHb W=0.094 8H2.821 0.951 3 25.43 W=a(D2H)b W=0.432 7(D2H)0.692 0.957 4 21.69 W=aDbHc W=0.184 3D0.758H1.708 0.968 1 18.53 W=a(D3/H)b W=2.111 0(D3/H)0.716 0.894 3 42.66 -
利用最优整株生物量模型W=0.184 3D0.758H1.708计算出3个林场共筛选的4 791个小班的平均整株生物量,并研究13个特征变量对雪岭云杉生物量的影响,用随机森林法中的变量相对重要性来描述。从图4可知:影响雪岭云杉生物量的变量重要性排序依次为坡向、下木盖度、坡位、疏密度、海拔、坡度、土壤厚度、每公顷蓄积量、下木高度、平均年龄、郁闭度、平均胸径和平均树高。

图 4影响雪岭云杉生物量的变量相对重要性
Figure 4.Relative importance of variables affectingP. schrenkianabiomass
-
生物量分配是植物生殖与生存平衡的结果[21],是一个比率驱动过程(ratio-driven process),主要受植物的遗传特性(自身生长发育)和环境条件的影响[22]。大量研究表明:植株地上生物量占整株生物量的79%左右[23-24];本研究发现:雪岭云杉地上和地下生物量为10.16~1 475.17 和1.88~539.18 kg·株−1,分别占总生物量的64.5%~92.4%和7.7%~35.5%。张绘芳等[25]发现阿尔泰山西伯利亚云杉干、枝、叶和根生物量分别占整株生物量的48.1%、20.0%、11.7%和20.2%,与本研究存在一定差异,主要是树种差异(树龄、种类、植株大小,环境因子中水分、温度、光照)和生长地域差异(坡度、土壤厚度等)对植物各器官生物量的分配造成影响,因此,后续研究若需调用某一乔木生物量公式,不适合直接套用不同区域同一树种的生长模型,否则可能对计算结果带来较大的误差。
大部分研究表明:植物的地下与地上生物量多存在线性关系[26]。本研究中雪岭云杉地上生物量与地下生物量呈幂函数关系,但相关性分析发现,两者为极显著正相关(R2
=0.918 2,P<0.001),即两者随着树木生长生物量共同增加。说明随着植株光合作用产物的积累,地上生物量逐渐增加,地下根系为支撑地上各器官部分的稳定必须获取更多的养分而逐步生长,即根系生物量也同步增加。植株地上生物量和地下生物量分配系数的相对稳定,是植物保持优势生长的必要条件。 -
本研究分别以D、H、D2H、D3/H和DbHc为自变量,构建雪岭云杉干、枝、叶、根、地上生物量及整株生物量的异速生长模型。筛选发现,树根最优生物量模型以D2H为自变量,其他各器官生物量最优模型以DbHc为自变量。与张绘芳等[25]的研究存在差异,可能是受实际调查客观条件、测量方法和立地条件的影响,相同树种不同环境、不同标准木胸径等也会导致异速生长模型的差异。因此,今后研究中,要注意对不同林龄及不同径阶下的云杉生物量分配模式及异速生长模型作出区分。
本研究建立的雪岭云杉异速生长模型,以“胸径+树高”为变量,拟合结果均优于“树高”和“胸径”单独建立的模型;白志强等[15]、张绘芳等[25]等认为受光照、水分、竞争和立地环境条件等非生物因素和某些生物因素的影响[27],雪岭云杉枝、叶生物量模型的解释率相对较低,因此在建立枝、叶生物量异速生长模型时,还需考虑东西南北冠幅、林龄、树冠长度等因素。
-
利用随机森林法对影响雪岭云杉生物量的变量重要性进行排序,发现坡向、坡位、坡度、海拔及土壤厚度等环境因子排序较靠前,与曾斌等[28]和单长卷[29]结论一致,表明不同环境因子组合对树种生长表现出不同程度的作用。因此,应客观考虑各环境因子的影响程度,使林木的生长环境条件尽可能地处于最佳组合状态[26,30]。
雪岭云杉多生长在海拔1 400~2 700 m的中山带阴坡;随海拔增加,光照强度增强,水分减少,雪岭云杉的生长受到制约,因此海拔越高,雪岭云杉分布也越稀疏。同时,雪岭云杉为喜阴植物,相对于阳坡,阴坡的长势更好;分析原因可知:阴坡接受太阳辐射少,土壤的水分蒸发慢,水土保持较好,土壤中物理、化学和生物过程的差异较明显,也更容易形成森林,而不同坡位的土壤水分、养分也存在着较大差异,对林木生长有着较大影响。因此,在人工林造林时,需考虑树种类型及树种适宜生存的环境等。
Biomass allocation and allometric growth ofPicea schrenkianain Tianshan Mountains
-
摘要:
目的雪岭云杉 Picea schrenkiana是新疆山区重要树种。了解雪岭云杉地上地下生物量分配及碳储量,对新疆森林资源调查具有一定意义。 方法采用整株收获法分析30株雪岭云杉地上地下生物量分配格局,利用胸径( D)、树高( H)和胸径-树高( D 2 H、 D 3/ H和 D b H c)作为变量建立树干、树枝、树叶、树根、地上及整株生物量异速生长模型。 结果雪岭云杉树干、树枝、树叶及树根生物量存在显著性差异( P<0.01)。整株生物量为12.04~2 014.34 kg·株 −1,地上和地下生物量分别为10.16~1 475.17和1.88~539.18 kg·株 −1,树干、树枝、树叶及树根生物量占整株生物量的56.86%、13.03%、5.96%和24.15%,根冠比为0.08~0.55。植株水平上,建立基于胸径及树高变量的各器官生物量模型,其中树根生物量的最优生物量模型为 W= a( D 2 H) b,其他器官生物量模型均为 W= aDbHc。影响云杉生物量的主要环境因素重要性排序依次为坡位、坡度、海拔及土壤厚度。 结论基于胸径-树高因素的异速生长模型可以较好地实现雪岭云杉各器官生物量的拟合,可对其生物量及碳储量进行有效估算。图4表3参30 Abstract:ObjectiveA proper understanding of the above-ground and underground biomass allocation and carbon storage of Picea schrenkiana, an important tree species in Xinjiang mountainous areas of great significance to the forest resources investigations in Xinjiang. To establish the allometric growth models of trunk, branch, leaf, root, the above-ground part and the whole plant with DBH( D), tree height ( H) and DBH-H ( D 2 H, D 3/ Hand DbHc). MethodAnalysis of the of above-ground and underground biomass distribution pattern of 30 P. schrenkianatrees by whole plant harvesting method. ResultThere were significant differences in the biomass of trunk, branch, leaf and root of P. schrenkiana( P<0.01). The biomass of whole P. schrenkianatrees ranged from 12.04−2 014.34 kg·plant −1, and the biomass of the above-ground part and the underground part were 10.16−1 475.17 and 1.88−539.18 kg·plant −1respectively. The biomass of trunk, branch, leaf and root accounted for 56.86%, 13.03%, 5.96% and 24.15% of the whole plant, and the range of root-shoot ratio was 0.08−0.55. At the plant level, the biomass model of each organ based on DBH( D) and tree height( H) variables was established. The optimal biomass model of root biomass was W= a( D 2 H) b, and other organs was W= aDbHc. Slope position, slope, altitude and soil thickness are the main environmental factors affecting the biomass of spruce. ConclusionThe allometric growth model based on tree height-DBH can better fit the biomass of each organ of P. schrenkiana, and can effectively estimate its biomass and carbon reserves. [Ch, 4 fig. 3 tab. 30 ref.] -
Key words:
- forest mensuration/
- biomass/
- allocation pattern/
- allometric model/
- organ/
- Picea schrenkiana
-
图 1雪岭云杉地上与地下生物量的关系
Figure 1Relationship between aboveground and underground biomass ofP. schrenkiana
图 2雪岭云杉不同器官生物量相对分配比例
Figure 2Relative allocation ratio of biomass in different organs ofP. schrenkiana
图 3天山雪岭云杉单木各器官生物量随径级变化图
Figure 3Variation of individual organ biomass with diameter ofP. schrenkianain Tianshan Mountains
图 4影响雪岭云杉生物量的变量相对重要性
Figure 4Relative importance of variables affectingP. schrenkianabiomass
表 1定性指标量化标准
Table 1.Qualitative indicator quantitative standard
评价标准 量化标准 评价标准 量化标准 坡向 坡位 土壤厚度/cm 坡向 坡位 土壤厚度/cm 无坡向 − − 9 东南 下 − 4 西北 − − 8 东 中 <30(薄) 3 西 − − 7 东北 上 30~59(中) 2 西南 平地 − 6 北 脊 ≥60(厚) 1 南 谷 − 5 说明:“−”表示无相关定性等级  下载:
导出CSV
下载:
导出CSV
表 2天山雪岭云杉单木各器官生物量特征表
Table 2.Characteristic table of individual organ biomass ofP. schrenkianain Tianshan Mountains
径级/cm 整株生物量/(kg·株−1) 地上生物量/(kg·株−1) 地下生物量/(kg·株−1) 根冠比 区间 平均 区间 平均 区间 平均 区间 平均 6.5~20.0 12.04~183.49 70.29 10.16~169.45 55.90 1.88~36.14 14.39 0.08~0.51 0.31 21.0~38.0 124.43~541.62 338.03 99.78~400.49 254.29 24.65~141.12 83.74 0.21~0.54 0.33 41.0~60.0 552.61~1 395.75 918.82 441.28~1 101.87 687.17 111.33~376.77 231.65 0.18~0.55 0.34 61.0~81.0 1 830.42~2 014.34 1 922.38 1 455.44~1 475.17 1 465.31 374.98~539.18 457.08 0.26~0.37 0.31 下载:
导出CSV
表 3新疆天山雪岭云杉各器官生物量5种非线性生物量估测模型
Table 3.Five nonlinear biomass estimation models ofP. schrenkianaorgans in Tianshan forest areas
器官 生物量模型 拟合公式 R2 EMP/% 树干 W=aDb W=0.476 6D1.803 0.926 6 38.98 W=aHb W=0.050 6H2.859 0.951 9 25.19 W=a(D2H)b W=0.251 7(D2H)0.695 0.947 5 30.69 W=aDbHc W=0.088 5D0.625H1.938 0.963 1 23.53 W=a(D3/H)b W=1.389 0(D3/H)0.706 0.876 7 63.69 树枝 W=aDb W=0.073 6D1.835 0.768 0 34.49 W=aHb W=0.003 8H3.114 0.830 7 43.52 W=a(D2H)b W=0.033 8(D2H)0.719 0.796 0 35.15 W=aDbHc W=0.004 9D0.252H2.736 0.832 5 42.78 W=a(D3/H)b W=0.139 0(D3/H)0.769 0.709 9 36.41 树叶 W=aDb W=0.167 2D1.395 0.753 6 40.68 W=aHb W=0.030 5H2.205 0.800 9 31.27 W=a(D2H)b W=0.100 9(D2H)0.539 0.777 1 35.39 W=aDbHc W=0.035 8D0.229H1.881 0.802 8 30.69 W=a(D3/H)b W=0.281 5(D3/H)0.581 0.687 9 47.56 树根 W=aDb W=0.178 8D1.826 0.904 4 37.43 W=aHb W=0.032 3H2.722 0.857 8 36.64 W=a(D2H)b W=0.100 6(D2H)0.697 0.908 7 31.85 W=aDbHc W=0.101 9D1.405H0.681 0.908 7 31.92 W=a(D3/H)b W=0.422 7(D3/H)0.741 0.880 0 53.82 地上生物量 W=aDb W=0.635 8D1.786 0.915 7 29.52 W=aHb W=0.064 1H2.854 0.948 4 24.17 W=a(D2H)b W=0.332 3(D2H)0.69 0.938 3 22.83 W=aDbHc W=0.104 8D0.551H2.042 0.957 3 19.09 W=a(D3/H)b W=1.701 0(D3/H)0.708 0.860 3 46.40 总生物量 W=aDb W=0.813 0D1.796 0.938 8 27.08 W=aHb W=0.094 8H2.821 0.951 3 25.43 W=a(D2H)b W=0.432 7(D2H)0.692 0.957 4 21.69 W=aDbHc W=0.184 3D0.758H1.708 0.968 1 18.53 W=a(D3/H)b W=2.111 0(D3/H)0.716 0.894 3 42.66 下载:
导出CSV
-
[1] 刘坤,曹林,汪贵斌,等. 银杏生物量分配格局及异速生长模型[J]. 北京林业大学学报, 2017,39(4): 12 − 20.LIU Kun, CAO Lin, WANG Guibin,et al. Biomass allocation patterns and allometric models ofGinkgo biloba[J].J Beijing For Univ, 2017,39(4): 12 − 20. [2] OLSON J S, WATTS J A, ALLISON L J.Carbon in Live Vegetation of Major World Ecosystems[R]. Oak Ridge: Oak Ridge Laboratory, 1983. [3] 方精云,刘国华,徐嵩龄. 我国森林植被的生物量和净生产量[J]. 生态学报, 1996,16(5): 497 − 508.FANG Jingyun, LIU Guohua, XU Songling. Biomass and net production of forest vegetation in China [J].Acta Ecol Sin, 1996,16(5): 497 − 508. [4] 邱东,周桂玲,刘同业. 3种棒果芥属植物生物量分配及异速生长分析[J]. 干旱地区农业研究, 2014,32(6): 215 − 220.QIU Dong, ZHOU Guiling, LIU Tongye. Analysis of biomass allocation and allometric growth of threeSterigmostemumspecies in Junggar Basin [J].Agric Res Arid Areas, 2014,32(6): 215 − 220. [5] 董道瑞,李霞,万红梅,等. 塔里木河下游胡杨(Populus euphratica)地上生物量估测[J]. 中国沙漠, 2013,33(3): 724 − 730.DONG Daorui, LI Xia, WAN Hongmei,et al. Aboveground biomass estimation ofPopulus euphraticain the lower reaches of Tarim River [J].J Desert Res, 2013,33(3): 724 − 730. [6] 汪金松,张春雨,范秀华,等. 臭冷杉生物量分配格局及异速生长模型[J]. 生态学报, 2011,31(14): 3918 − 3927.WANG Jinsong, ZHANG Chunyu, FAN Xiuhua,et al. Biomass allocation patterns and allometric models ofAbies nephrolepisMaxim [J].Acta Ecol Sin, 2011,31(14): 3918 − 3927. [7] 于贵瑞. 全球变化与陆地生态系统碳循环和碳蓄积[M]. 北京: 气象出版社, 2003: 157 − 158. [8] 董鸣. 陆地生物群落调查观测与分析[M]. 北京: 中国标准出版社, 1997: 73 − 75. [9] 刘琪璟. 嵌套式回归建立树木生物量模型[J]. 植物生态学报, 2009,33(2): 331 − 337.LIU Qijing. Nested regression for establishing tree biomass equations [J].Chin J Plant Ecol, 2009,33(2): 331 − 337. [10] 张志,田昕,陈尔学,等. 森林地上生物量估测方法研究综述[J]. 北京林业大学学报, 2011,33(5): 144 − 150.ZHANG Zhi, TIAN Xin, CHEN Erxue,et al. Review of methods on estimating forest above ground biomass [J].J Beijing For Univ, 2011,33(5): 144 − 150. [11] NIKLAS K J.Plant Allometry:The Scaling of Form and Process[M]. Chicago: University of Chicago Press, 1994. [12] 朱江,韩海荣,康峰峰,等. 山西太岳山华北落叶松生物量分配格局与异速生长模型[J]. 生态学杂志, 2016,35(11): 2918 − 2925.ZHU Jiang, HAN Hairong, KANG Fengfeng,et al. Biomass allocation patterns and allometric models ofLarix principis-rupprechtiiin Mt. Taiyue, Shanxi [J].Chin J Ecol, 2016,35(11): 2918 − 2925. [13] 汪珍川,杜虎,宋同清,等. 广西主要树种(组)异速生长模型及森林生物量特征[J]. 生态学报, 2015,35(13): 4462 − 4472.WANG Zhenchuan, DU Hu, SONG Tongqing,et al. Allometric models of major tree species and forest biomass in Guangxi [J].Acta Ecol Sin, 2015,35(13): 4462 − 4472. [14] 张绘芳,朱雅丽,地力夏提•包尔汉,等. 阿尔泰山林区云杉和落叶松生物量分配格局研究[J]. 南京林业大学学报(自然科学版), 2017,41(1): 203 − 208.ZHANG Huifang, ZHU Yali, Dilixiati Baoerhan,et al. Biomass allocation patterns ofPicea obovataandLarix sibiricain the Altai Mountains forest area [J].J Nanjing For Univ Nat Sci Ed, 2017,41(1): 203 − 208. [15] 白志强,李缓,王文栋,等. 阿尔泰山优势树种的生物量模型构建及其生物量分配特征[J]. 林业资源管理, 2018(4): 34 − 40.BAI Zhiqiang, LI Huan, WANG Wendong,et al. Biomass model establishment and allocation of dominant tree species in Altai Mountains [J].For Res Manage, 2018(4): 34 − 40. [16] 孟宪宇. 测树学[M]. 北京: 中国林业出版社, 2006: 197-199. [17] 明安刚,郑路,麻静,等. 铁力木人工林生物量与碳储量及其分配特征[J]. 北京林业大学学报, 2015,37(2): 32 − 39.MING Angang, ZHENG Lu, MA Jing,et al. Biomass, carbon stock and allocation characteristics inMesua ferreaplantation [J].J Beijing For Univ, 2015,37(2): 32 − 39. [18] GOWER S T, KUCHARIK C J, NORMAN J M. Direct and indirect estimation of leaf area index,fAPAR, and net primary production of terrestrial ecosystems [J].Remote Sensing Environ, 1999,70(1): 29 − 51. [19] CASE B S, HALL R J. Assessing prediction errors of generalized tree biomass and volume equations for the boreal forest region of west-central Canada [J].Can J For Res, 2008,38(6): 878 − 889. [20] 国家林业局. 国家森林资源连续清查技术规定[R]. 北京: 国家林业局, 2014. [21] 郝虎东,田青松,石凤翎,等. 无芒雀麦地上生物量及各构件生物量分配动态[J]. 中国草地学报, 2009,31(4): 85 − 90.HAO Hudong, TIAN Qingsong, SHI Fengling,et al. Allocated dynamics of aboveground biomass and structural biomass inBromus inermisLeyss [J].Chin J Grassland, 2009,31(4): 85 − 90. [22] 李涛. 荒漠植物异速生长指数及其空间格局适应降雨梯度变化的规律与机制研究[D]. 兰州: 兰州大学, 2010.LI Tao.Response Mechanism of Desert Plant Allometric Exponents and Spatial Patterns to A Precipitation Gradient[D]. Lanzhou: Lanzhou University, 2010. [23] HOUGHTON R A, LAWRENCE K T, HACKLER J L,et al. The spatial distribution of forest biomass in the Brazilian Amazon: a comparison of estimates [J].Global Change Biol, 2001,7: 731 − 746. [24] SAATCHI S S, HOUGHTON R A, DOS SANTOS ALVALA R C,et al. Distribution of aboveground live biomass in the Amazon Basin [J].Global Change Biol, 2007,13(4): 816 − 837. [25] 张绘芳,高亚琪,朱雅丽,等. 新疆雪岭杉生物量模型对比研究[J]. 西北林学院学报, 2015,30(6): 52 − 58.ZHANG Huifang, GAO Yaqi, ZHU Yali,et al. A comparative study on biomass models forPicea schrenkianain Xinjiang [J].J Northwest For Univ, 2015,30(6): 52 − 58. [26] BROWN S. Measuring carbon in forests: current status and future challenges [J].Environ Pollut, 2002,116(3): 363 − 372. [27] BOND-LAMBERTY B, WANG C, GOWER S T. Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba [J].Can J For Res, 2002,32: 1441 − 1450. [28] 曾斌,翟学昌,彭丽,等. 赣南不同立地杉木人工林生产力研究[J]. 安徽农业科学, 2010,38(14): 7625 − 7626.ZENG Bin, ZHAI Xuechang, PENG Li,et al. Study on productivity of fir plantation in different sites in southern Jiangxi [J].J Anhui Agric Sci, 2010,38(14): 7625 − 7626. [29] 单长卷. 黄土高原不同立地刺槐林水分关系研究[D]. 杨凌: 西北农林科技大学, 2004.SHAN Changjuan.Study on the Relationship Between Soil Water and Water Eco-Physiological Characteristics of Locust in Different Habitas of the Loess Plateau[D]. Yangling: Northwest A&F University, 2004. [30] 赖文豪,席沁,武海龙,等. 内蒙古兴和县低山丘陵立地类型划分与林草适宜性评价[J]. 浙江农林大学学报, 2018,35(2): 331 − 339.LAI Wenhao, XI Qin, WU Hailong,et al. Site classification type and vegetation suitabity for hilly land in Xinghe, Inner Mongolia [J].J Zhejiang A&F Univ, 2018,35(2): 331 − 339. -

- 链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190384
 点击查看大图
点击查看大图

计量
- 文章访问数:4628
- HTML全文浏览量:1455
- PDF下载量:147
- 被引次数:0











