




 下载:
下载:







-
苯丙氨酸解氨酶(PAL)是植物次生代谢途径中的关键酶,催化苯丙氨酸转化成反式肉桂酸[1],进而通过苯丙烷代谢途径调控木质素、黄酮类、香豆素等化合物的合成,对植物生长发育和环境适应至关重要。PAL基因家族成员在不同植物中的数量差异显著,如拟南芥Arabidopsis thaliana[2]和烟草Nicotiana tabacum各有4个[3],水稻Oryza sativa有9个[4],核桃Juglans regia有12个[5],这些基因可能具有不同的表达模式和功能冗余。拟南芥AtPAL1和AtPAL2影响花青素和原花青素的合成,且存在功能冗余[6],而四重突变体(pal1/2/3/4)表现出生长和发育缺陷[2]。在梨Pyrus bretschneideri中,PbPAL1影响石细胞和木质素合成[7]。此外,PAL基因在植物防御反应中也发挥重要作用。火炬松Pinus taeda的PAL在生物和非生物胁迫下的表达模式,表明其参与植物防御反应[8]。高粱Sorghum bicolor蚜虫侵害的研究发现:甘蔗蚜虫诱导SbPAL基因表达,通过正向调控水杨酸介导的防御信号通路抑制蚜虫种群的生长和繁殖[9]。由此可见,深入研究PAL基因家族的功能,对于提升植物生长、抗病抗虫能力以及次生代谢物产量具有重要意义。
山核桃Carya cathayensis为胡桃科Juglandaceae山核桃属Carya落叶乔木,种植历史悠久[10]。山核桃次生代谢物丰富,具有重要的营养价值和观赏价值,果实为世界四大名优坚果之一[11]。然而,在离体培养过程中,山核桃由于酚类物质如黄酮类、木质素及单宁等大量积累,存在严重的褐化问题[12]。PAL作为调控这些酚类物质合成的关键酶,其结合位点和表达模式对控制致褐物质合成至关重要。通过对欧芹Petroselinum crispum中苯丙氨酸解氨酶(PcPAL)的疏水结合口袋进行改造,可以提高PcPAL对苯丙氨酸和肉桂酸单取代衍生物的活性和选择性[13]。而将PcPAL的底物特异性调节剂残基L134、F137、I460印记到拟南芥AtPAL和多变鱼腥藻Anabaena variabilis的AvPAL上,可揭示不同PALs在底物特异性上的差异,并为底物特异性的调节提供更多信息[14]。通过研究不同PAL成员的结构功能和底物偏好,找到控制褐化物质合成的关键PAL基因,对于山核桃再生体系和遗传转化体系的建立具有重要作用。本研究基于山核桃全基因组测序和转录组测序的结果,筛选山核桃PAL(CcPAL)基因家族成员,研究CcPAL编码蛋白的理化性质和结构特点,为研究CcPAL在山核桃酚类物质合成以及愈伤组织增殖与褐化、组织分化和器官再生方面的作用机制提供参考。
-
山核桃愈伤组织由浙江农林大学玲珑山山核桃试验林#34山核桃树的未成熟合子胚诱导而来,果实采集时间为2021年7月中上旬。跟踪山核桃愈伤组织增殖褐化过程,选择了继代后8个时间点(继代后0、3、7、10、14、17、21和24 d)的愈伤组织用于转录组测序。
-
为鉴定CcPAL基因家族成员,从拟南芥信息资源中心数据库(TAIR)(www.arabidopsis.org)中获取4条AtPAL蛋白序列,在美国国家生物技术信息中心(NCBI)获得JrPAL(JX069977.1)[15]基因序列,分别作为query序列检索山核桃PAL成员,进行交叉验证。通过TBtools软件[16]在山核桃基因组数据[17]和转录组数据中进行BLAST比对,设定比对期望值E<10−10,筛选山核桃PAL同源序列。通过InterPro (www.ebi.ac.uk/interpro)网站[18]确认PAL蛋白的特征结构域PF00221,进一步验证候选基因家族成员。根据山核桃染色体组装结果,将各成员依次命名为CcPAL1~CcPAL6。
-
利用ProtParam (
https://web.expasy.org/protparam/ )网站[19]在线预测CcPALs蛋白的氨基酸长度、相对分子量、等电点、脂肪系数及总平均亲水系数等理化性质。同时通过DeepLoc2.1 (https://services.healthtech.dtu.dk/services/DeepLoc-2.1/ )网站[20]对CcPAL家族蛋白的亚细胞分布位置进行预测分析。 -
通过SOPMA (
https://npsa-prabi.ibcp.fr/ )网站[21]和Phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index )网站[22]对CcPALs的二级结构和三级结构进行预测分析和可视化。 -
利用山核桃基因组注释文件,使用TBtools软件Visualize Gene Structure模块可视化CcPALs基因结构图;通过NCBI网站中的Batch CD-Search工具(
https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )获得CcPALs蛋白保守结构域,使用TBtools软件Visualize NCBI CDD Domain Pattern模块可视化结构域;通过MEME (https://meme-suite.org )网站[23]分析CcPALs蛋白的保守基序,并使用TBtools软件Visualize MEME/MAST Motif Pattern模块绘制图像。 -
利用MEGA 11[24]对山核桃、拟南芥、水稻Oryza sativa、毛果杨Populus trichocarpa、核桃、杉木Cunninghamia lanceolata、海岸松Pinus pinaster和火炬松Pinus taeda等8个物种42条PAL蛋白序列进行比对,采用邻接法(NJ),步长设为1 000,其余参数默认,构建系统进化树。
-
收集#34山核桃树继代后0、3、7、10、14、17、21和24 d (对应A、B、C、D、E、F、G和H样本)的愈伤组织样品,每个时间点3个重复。样品冻存于−80 ℃冰箱,研磨粉碎,参考MonPure™ Plant RNA Kit (polysaccharide & polyphenol-rich)(Monad,cat.MI10101S)高多糖多酚RNA提取试剂盒产品说明书的方法,提取山核桃愈伤总RNA。质检合格后委托杭州联川生物技术股份有限公司构建cDNA文库,对24个样品RNA进行有参转录组分析。由杭州联川生物技术股份公司进行下机原始数据的质控,浙江农林大学提供山核桃基因组数据[17]进行测序比对,之后对基因或转录本进行组装、每千个碱基的转录每百万映射读取的片段(FPKM)定量以及差异基因分析。
-
通过联川生物云平台(https://www.omicstudio.cn/home)分析山核桃愈伤组织的CcPALs基因表达量,并制作热图。
选择继代后0、7、14和21 d山核桃愈伤组织进行实时荧光定量PCR (RT-qPCR)表达量验证。采用多糖多酚RNA提取试剂盒(DR0407050,浙江易思得生物科技有限公司)分别提取愈伤组织的总RNA。通过反转录试剂盒(RR036A,北京宝日医生物技术有限公司)分别合成cDNA,置于−80 ℃冰箱备用。将愈伤组织的cDNA质量浓度调整到100 ng·μL−1,采用NovoStart®SYBR qPCR SuperMix Plus(E096,苏州近岸蛋白质科技股份有限公司)在CFX96 Touch荧光定量PCR仪(BIO-RAD,美国)上检测CcPALs基因的表达情况。β-actin为内参基因,引物信息见表1,每个样品3个生物学重复,通过GraphPad Prism软件进行单因素方差分析(one-way ANOVA)和制图。
表 1实验中使用的RT-qPCR引物
Table 1.RT-qPCR primers used in the experiment
名称 上游引物(5′→3′) 下游引物(5′→3′) β-actin ATGGCCGATGCTGAGGATATT CACCAGTGTGTCGAGGTCTGC CcPAL1 ATGGGTTGCCTTCGAATCTC CCAGGGAGTTCACATCTTGG CcPAL2 AGGTTACTCGGGCATCAGAT GCCTATCAAAAGACCAGCGA CcPAL3 CTGCAGCGAAACCTATCCAT TGGCAGTGTTCAAGTTCCTC CcPAL4 CAGTGCGACATACCCATTGA TTCCTCAAAGACCCCGATCT CcPAL5 GCTTGGGGTAGAGGTAGCAA GCATTGGAGAAGTCACCGTG CcPAL6 TCAGGGGATCTTGTACCGTT GCAGAAATCCCAGCTCTTGT -
通过生物信息学分析发现:无论是以拟南芥AtPALs作为query序列检索,还是以亲缘关系更近的核桃JrPALs进行检索,都是鉴定到6个CcPALs蛋白,分别命名为CcPAL1 (CCA0546S0150)、CcPAL2 (CCA0589S0228)、CcPAL3 (CCA0589S0229)、CcPAL4 (CCA1013S0012)、CcPAL5 (CCA1591S0016)和CcPAL6 (CCA1591S0017)。分析CcPAL家族成员的氨基酸组成和理化性质发现:CcPALs蛋白含有的氨基酸数量为657~760个;6个CcPALs家族成员中亮氨酸(Leu)、丙氨酸(Ala)、甘氨酸(Gly)和谷氨酸(Glu)含量最为丰富;分子量为71.87~83.88 kDa;等电点为6.01~6.41,均小于7.00,属于酸性蛋白;所有CcPALs蛋白的不稳定系数均小于40,稳定性较高;6个CcPALs蛋白平均亲水性系数均小于0,均属于亲水蛋白,其中CcPAL6亲水性最强(表2)。
表 2CcPALs蛋白理化性质
Table 2.Physicochemical properties of CcPALs protein
蛋白质 氨基酸数量/个 主要氨基酸组分(相对占比/%) 理论等电点 分子量/kDa 不稳定系数 脂肪指数 平均亲水性系数 CcPAL1 713 Leu(10.7)、Ala(9.8)、Gly(7.7) 6.41 77.54 32.96 91.85 −0.149 CcPAL2 657 Leu(11.4)、Ala(9.1)、Glu(7.3) 6.07 71.87 32.62 95.65 −0.114 CcPAL3 708 Leu(11.0)、Ala(9.2)、Gly(7.5) 6.23 77.34 32.40 92.75 −0.189 CcPAL4 710 Leu(10.3)、Ala(9.9)、Glu(8.0) 6.10 77.50 37.69 91.56 −0.160 CcPAL5 712 Leu(10.1)、Ala(8.6)、Glu(7.7) 6.01 78.57 34.77 90.27 −0.174 CcPAL6 760 Leu(10.3)、Ala(8.8)、Glu(7.5) 6.22 83.88 35.75 89.08 −0.214 说明:Leu. 亮氨酸; Ala. 丙氨酸; Gly. 甘氨酸; Glu. 谷氨酸。 对CcPALs蛋白进行亚细胞定位预测发现:除CcPAL5定位在细胞核和内质网上外,CcPAL1、CcPAL2、CcPAL3、CcPAL4和CcPAL6可能在细胞核、线粒体和内质网均有定位(表3),说明CcPALs参与了基因转录调控、氧化磷酸化和蛋白质的加工等环节,在细胞生长周期中发挥着作用。
表 3CcPALs蛋白亚细胞定位预测
Table 3.Subcellular localization prediction of CcPALs protein inC. cathayensis
蛋白质 预测定位 定位信号 膜类型 CcPAL1 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL2 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL3 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL4 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL5 细胞核、内质网 核定位信号 外周、脂锚定 CcPAL6 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 对CcPALs的6个蛋白进行二级结构和三级结构预测分析发现:α-螺旋与无规则卷曲在二级结构中占比较高。其中CcPAL4的α-螺旋占比最高(56.48%),CcPAL5的延伸链结构、β-转角占比最高(9.13%、6.88%);CcPAL6的无规则卷曲占比最高(33.29%)(表4)。进一步对6个CcPAL蛋白进行了三级结构模型预测(图1),结果显示编码蛋白三级结构主要以α-螺旋为主要结构元件。
表 4CcPALs蛋白二级结构
Table 4.Secondary structure of CcPALs protein
蛋白质 占比/% α-螺旋 延伸链结构 β-转角 无规则卷曲 CcPAL1 53.02 8.70 6.73 31.56 CcPAL2 55.71 8.37 6.09 29.83 CcPAL3 55.37 7.49 6.50 30.65 CcPAL4 56.48 7.46 5.49 30.56 CcPAL5 55.34 9.13 6.88 28.65 CcPAL6 51.97 8.29 6.45 33.29 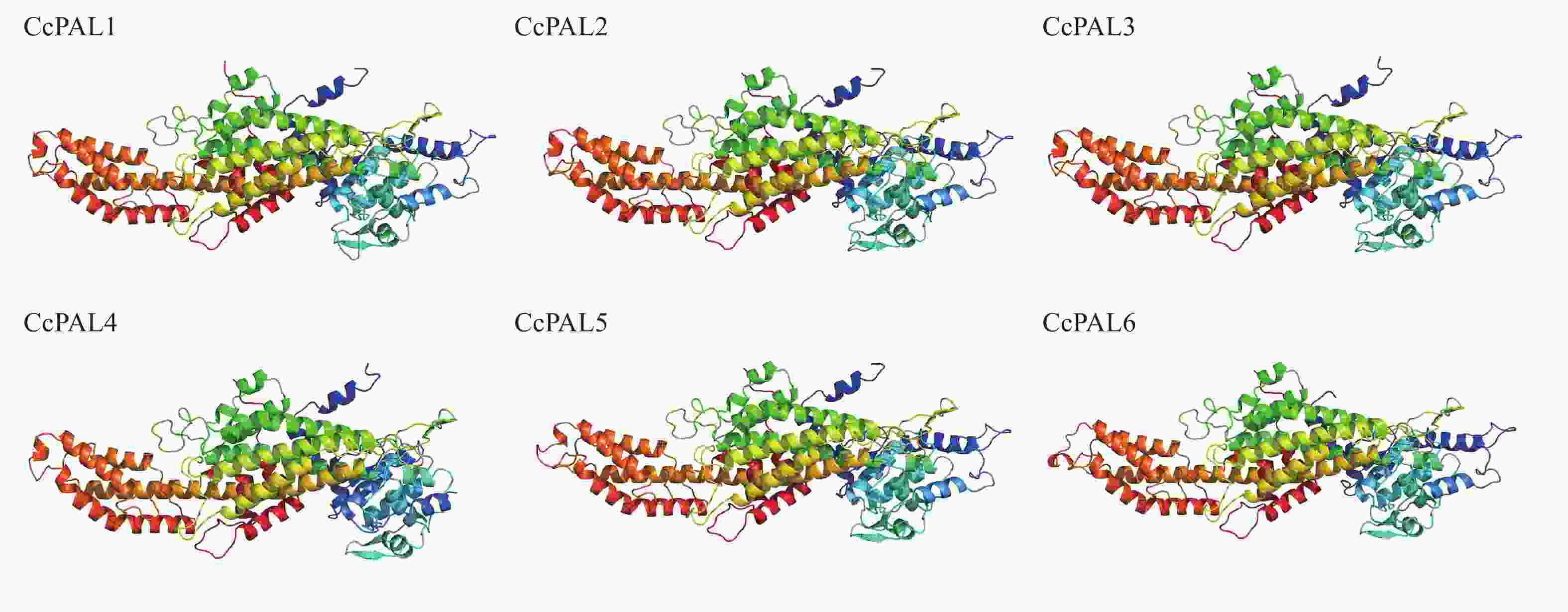
图 1CcPALs蛋白质三级结构
Figure 1.Tertiary structure of CcPALs protein
-
对CcPALs进行基因结构分析发现:CcPAL1~CcPAL4都各有2个外显子和1个内含子,结构相似,而CcPAL5和CcPAL6均只有1个外显子,且基因中不包含内含子序列(图2A)。6条CcPALs基因编码序列(CDS)长度为1 974~2 283 bp。

图 2CcPAL家族的基因结构(A)、保守蛋白(B)、保守基序(C)和基序标识分析(D)
Figure 2.Analysis of gene structure (A), conserved proteins (B), conserved motifs (C), and motif logos (D) of theCcPALfamily
通过NCBI的Conserved Domain Search对山核桃PAL家族氨基酸序列进行蛋白功能分析(图2B)显示:PLN02457 (phenylalanine ammoni-lyase)结构域在CcPALs中高度保守,它是Lyase_I_like超家族的一部分。对CcPAL家族蛋白进行保守基序分析,6个CcPALs蛋白都含有10个保守基序,且蛋白结构大体一致,说明CcPALs成员之间具有较高的保守性(图2C和图2D)。
-
MIO结构域在氨基酸的转化过程中通过消除氨形成不饱和产物,与PAL的催化活性密切相关。它包含PAL家族的保守序列(GTITASGDLVPLSYIAG)和丙氨酸-丝氨酸-甘氨酸(Ala-Ser-Gly)三肽活性中心。多重序列比对结果(图3)显示:CcPALs所有序列都含有MIO结构域(如CcPAL1的195~211个氨基酸)和Core Domain中的PAL活性位点(如CcPAL1上的348Tyr和491Gly),说明6个CcPALs都具有解氨酶催化活性。而MIO结构域在核桃JrPAL2、JrPAL3、JrPAL6和JrPAL9中完全缺失,在JrPAL8中缺失了4个氨基酸,且JrPAL1、JrPAL2、JrPAL3、JrPAL6、JrPAL7和JrPAL9与其他PAL在蛋白长度上表现出很大差异,缺乏明显的主要结构域PRK09367。另外Shielding Domain中的Thr残基被认为是潜在的磷酸化位点(CcPAL1中546Thr)。

图 3CcPAL蛋白氨基酸比对
Figure 3.Amino acid alignment of CcPAL protein
构建PAL基因家族系统进化树(图4)发现:PAL家族蛋白主要分成3个支。Group Ⅰ主要包含双子叶植物的PAL蛋白(用橙色表示),包括拟南芥AtPAL1~AtPAL4、毛果杨PtPAL1~PtPAL5,核桃JrPAL1、JrPAL2-2、JrPAL3、JrPAL5和JrPAL7以及山核桃CcPAL1~CcPAL4;Group Ⅱ则由单子叶植物的PAL蛋白组成(用绿色表示),如水稻OsPAL1~OsPAL9;Group Ⅲ由裸子植物PAL蛋白(海岸松PpPAL1和PpPAL2,火炬松PtaPAL1以及杉木ClPAL1~ClPAL3)和2条CcPAL (CcPAL5、CcPAL6)、7条JrPAL (JrPAL2-1、JrPAL2-3、JrPAL4-1、JrPAL4-2、JrPAL6、JrPAL8和JrPAL9)构成(用蓝色表示)。系统进化树分析结果显示:单子叶和双子叶植物的PAL家族成员在进化过程中呈现明显的分歧。山核桃的CcPAL5和CcPAL6与裸子植物的PAL聚在Group Ⅲ,表明在进化上这2个PAL蛋白与裸子植物同源性较高,功能上可能更相似。这一发现为进一步探索PAL基因家族的功能分化和进化提供了新的视角。
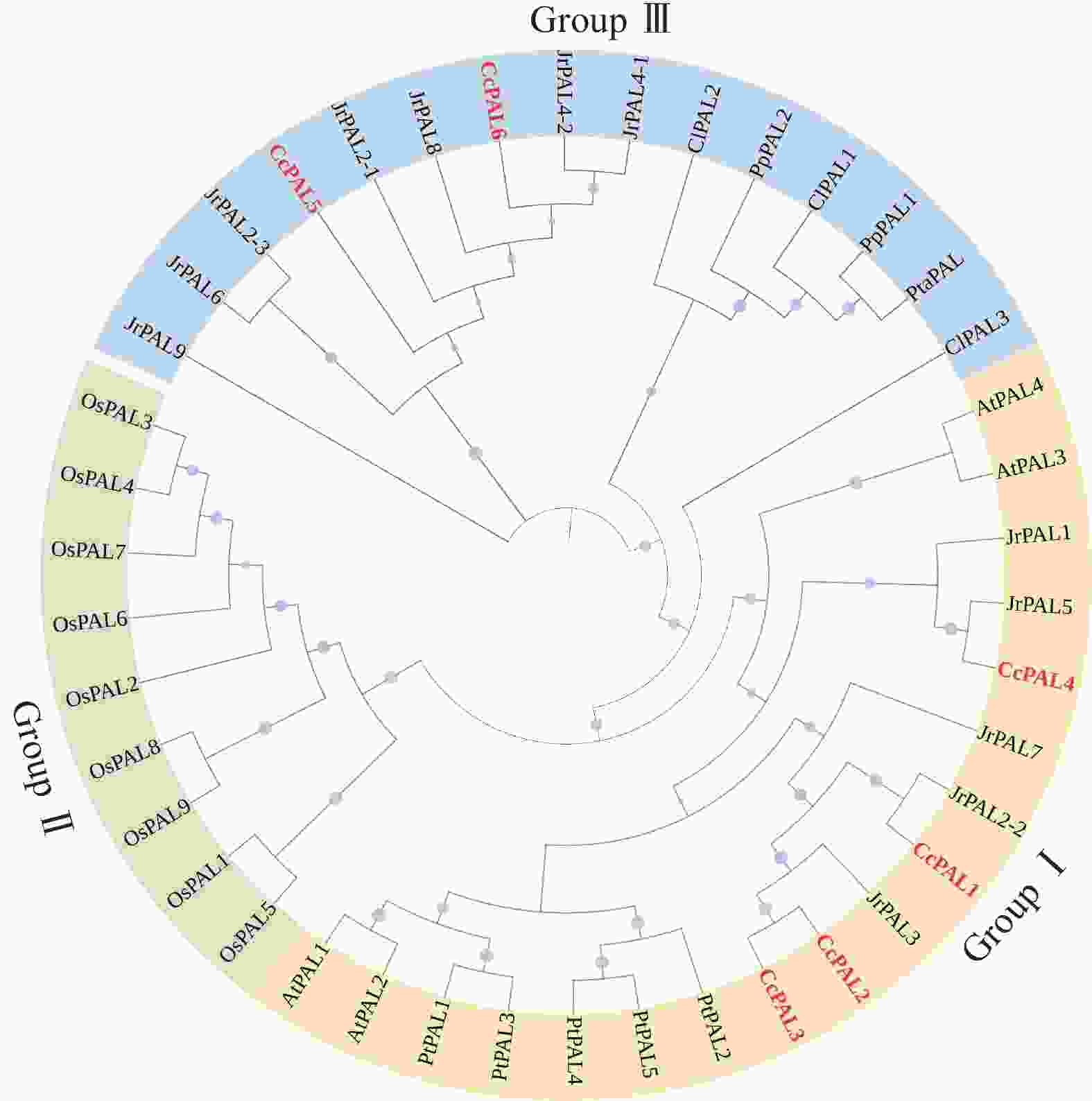
图 48个物种PAL家族蛋白的进化发育树
Figure 4.Phylogenetic tree of PAL family proteins in 8 species
-
基于Illumina NovaSeq6000测序平台构建文库。山核桃愈伤组织增殖褐化过程中8个时间点构建了24个cDNA文库。各样本Q30碱基百分比(碱基质量为30的碱基在总碱基数量中所占的比例)均高于96%,GC碱基占总碱基的44%~45%,符合正态分布。测序样本间转录本的主成分分析(PCA)显示:主成分1、主成分2的贡献率分别为74.11%和10.92%,每组3个样本间比较聚集,各自组内重复性较好,可用于后续分析(图5)。通过转录组测序,分析了山核桃愈伤组织在继代培养过程中从增殖到褐化阶段CcPALs基因的表达模式,并根据FPKM值绘制了热图(图6)。结果显示:聚类在同一分支的CcPALs表现出相似的表达趋势。继代7 d内,CcPAL1、CcPAL2、CcPAL3和CcPAL4表达量先上升后下降,CcPAL5和CcPAL6下调表达;继代14 d后除了CcPAL1和CcPAL4上调表达外,剩下4条CcPALs均下调表达;继代21 d后随着愈伤组织增殖减缓,出现褐化,CcPAL4先上调后下调,另外5条CcPALs先下调后上调;随着褐化的加重,6个CcPALs基因均上调表达。
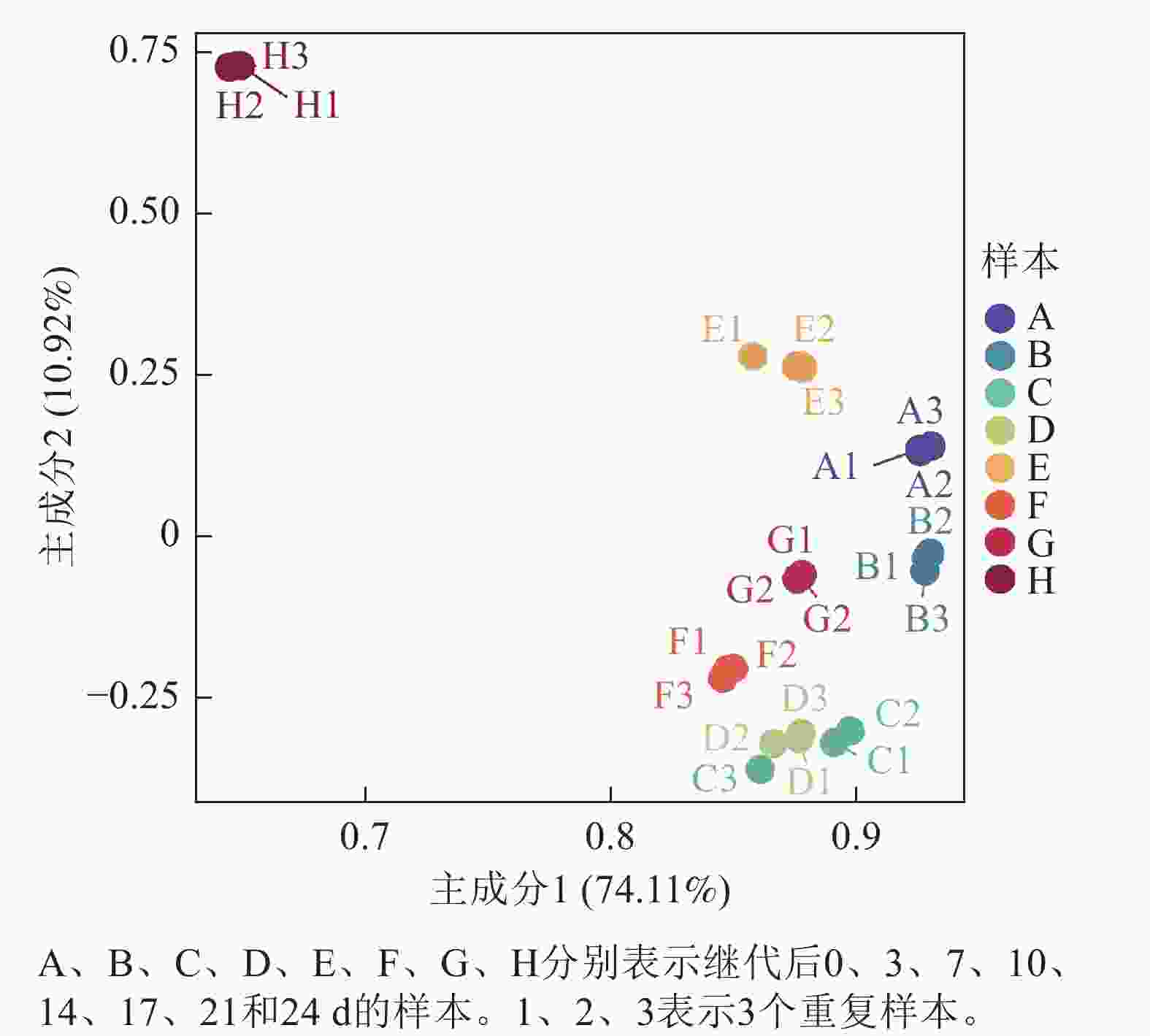
图 5测序样本间转录本PCA分析
Figure 5.PCA analysis of transcripts between sequencing samples
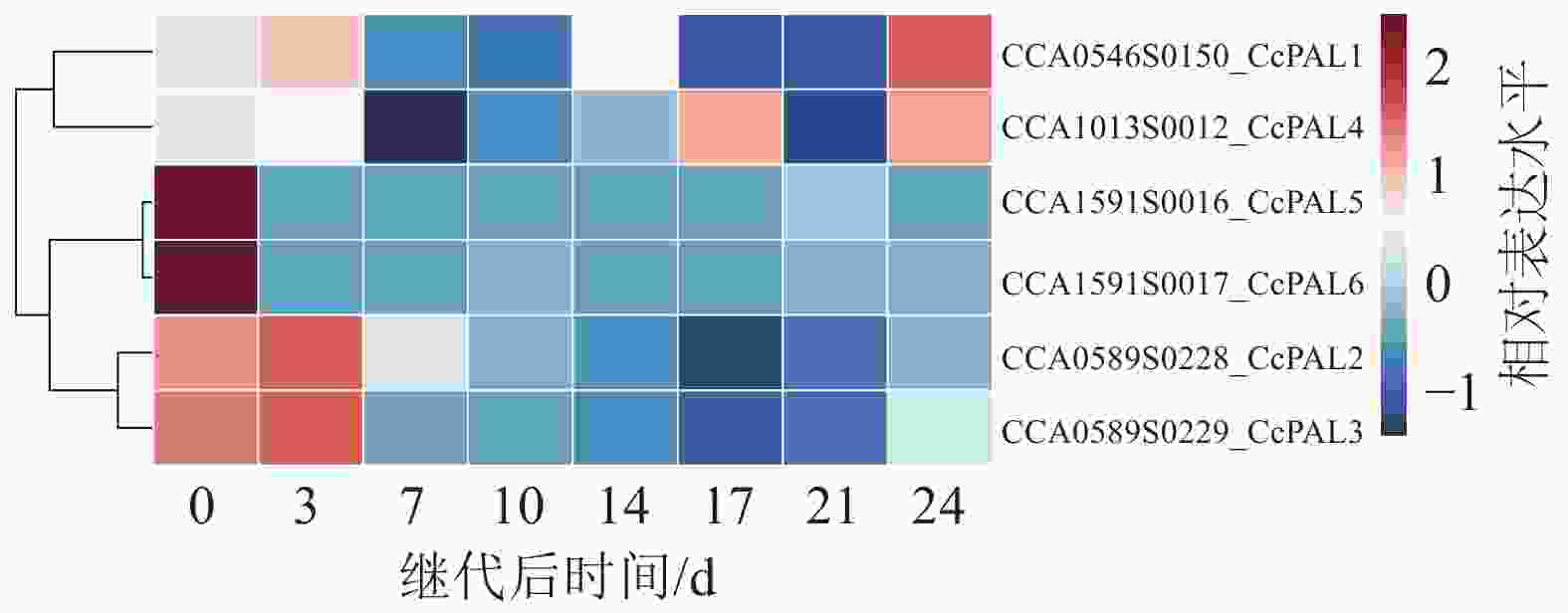
图 6山核桃愈伤组织增殖过程中CcPALs基因表达热图
Figure 6.Heat map ofCcPALsexpression during the proliferation of callus inC. cathayensis
CcPALs基因在不同时间的愈伤组织中均有表达,但不同基因的表达量不相同。CcPAL5和CcPAL6整体表达量较高,且在继代后0 d表达量最高,说明继代后愈伤组织受伤期间CcPAL5和CcPAL6已经开始响应表达;CcPAL1~CcPAL4基因在愈伤组织开始增殖(继代后7 d)和出现褐化(继代后21 d)时表达量较高,说明2个不同分支下的PAL基因对山核桃愈伤组织的环境适应和细胞增殖褐化有不同的影响(图7)。
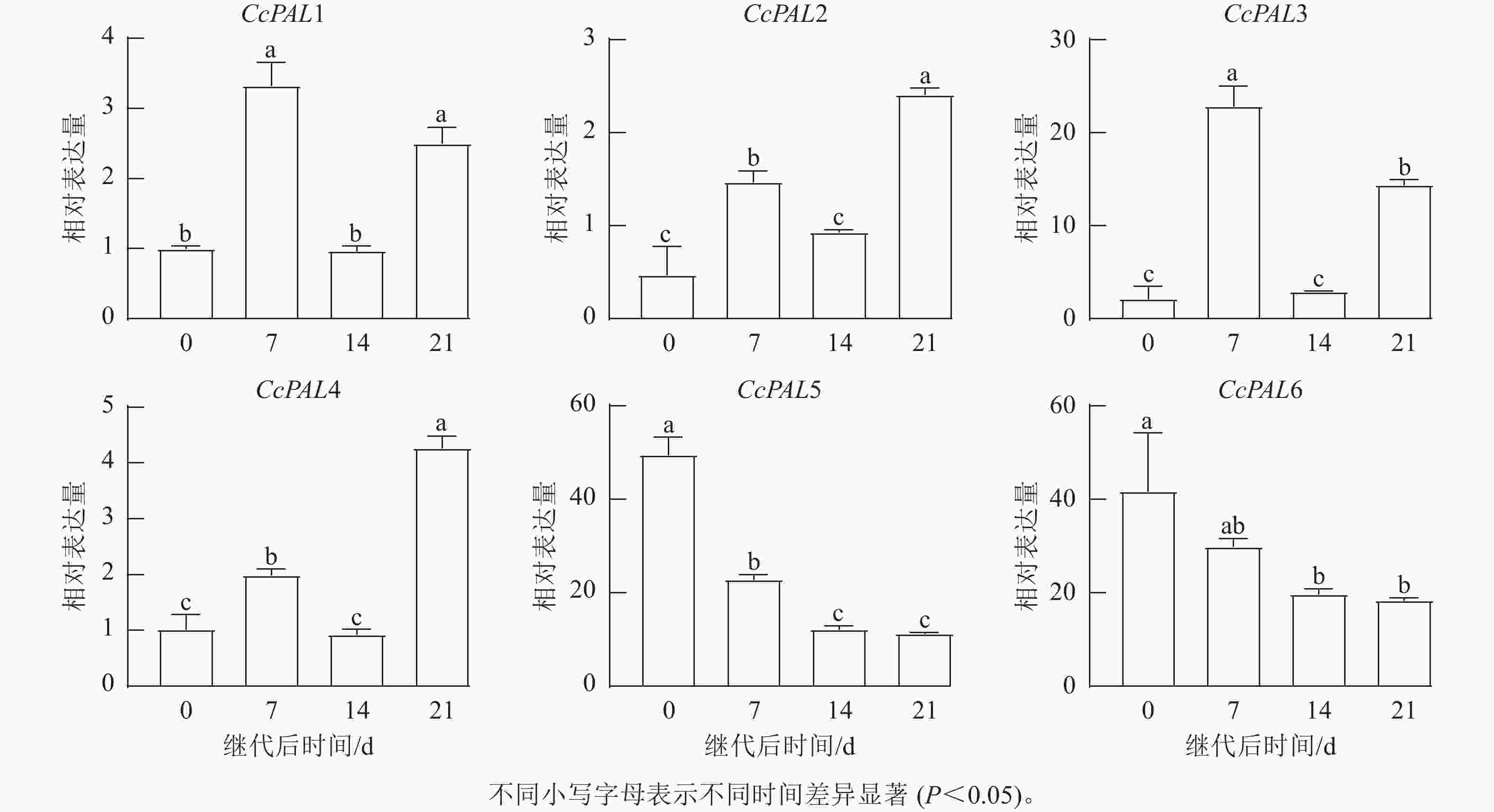
图 7继代后不同时间下山核桃愈伤组织中的CcPALs基因表达
Figure 7.Expression ofCcPALsgene in theC. cathayensiscallus at different time points after subculture
-
CcPALs蛋白的理化性质、基因结构分析结果与其他植物中的PAL蛋白具有一定的相似性和关联性[15,25−26]。在功能鉴定的PAL基因中都包含保守的GTITASGDLVPLSYIA特征基序、丙氨酸-丝氨酸-甘氨酸(Ala-Ser-Gly)三肽活性中心[27−28]和PAL活性位点氨基酸[29−30],而且在序列的屏蔽域中存在着磷酸化位点,即Thr残基[31]。所有CcPAL蛋白的不稳定系数小于40,属于稳定蛋白。这一特性对于山核桃在生长过程中维持PAL活性至关重要。在其他植物中,稳定的PAL蛋白也有助于植物稳定地参与次生代谢途径。例如,在火炬松中,PAL基因在植物防御反应中发挥着重要作用,其稳定性可能有助于火炬松在面对生物和非生物胁迫时保持代谢的稳定[8]。此外,CcPAL蛋白平均亲水性系数均小于0,均属于亲水蛋白。亲水性质可能使得这些蛋白更容易与其他分子相互作用,参与到各种代谢过程中。
基因结构方面,CcPAL1~CcPAL4与CcPAL5和CcPAL6的外显子和内含子结构不同。这种差异可能反映了不同的进化路径和分化功能。在拟南芥中,不同的PAL成员也可能具有不同的基因结构和表达模式[2,6]。例如,拟南芥中的PAL1和PAL2在黄酮类生物合成中有冗余作用,四重突变体表现出矮小和不育,这暗示着不同的PAL基因在功能上可能存在差异,而这种差异可能与基因结构的不同有关。CcPAL1~CcPAL4与CcPAL5和CcPAL6的基因结构差异可能也导致了它们在山核桃生长发育和代谢过程中的不同功能。
基因复制是进化创新和功能分化的主要机制[32]。核桃中存在12个PAL基因,彼此间蛋白结构差异较大,部分功能结构域缺失;而山核桃中存在6个PAL基因,成员间氨基酸数量相当,结构较为完整,存在个别氨基酸突变,说明PAL在核桃中发生了重大的变异和较多的基因复制活动,在山核桃中PAL更为保守。裸子植物的PAL基因亚群与被子植物的亚群是姐妹群,这可能意味着这些基因编码的PAL同工酶具有相似的功能,或者受到相似的发育调控机制的调控[8]。根据已测序物种的基因序列构建包含裸子植物、单子叶植物和双子叶植物的PAL系统发育树,结果显示:CcPAL5、CcPAL6和裸子植物PAL聚在一起,而CcPAL1~CcPAL4和双子叶植物聚在一起且具有高自展值,说明CcPAL5、CcPAL6与CcPAL1~CcPAL4可能有不同的进化起源。其他PAL基因在双子叶植物和单子叶植物分化后呈单系,表明基因复制在双子叶植物和单子叶植物分化后PAL基因的进化中很重要[33−35]。
山核桃愈伤组织增殖过程中不同的CcPALs基因表现出不同的表达模式,这与其他植物在生长发育或胁迫响应过程中PAL基因的差异表达现象类似[36]。在拟南芥中,PAL1和PAL2在黄酮类生物合成中有冗余作用,而本研究的山核桃中,CcPAL2和CcPAL3之间、CcPAL5和CcPAL6之间表达趋势相同,可能彼此间功能也存在冗余。在继代后3 d,CcPAL5和CcPAL6表达量显著上调,说明其可能参与机械损伤后抗逆相关黄酮类物质的合成,提高愈伤组织的环境适应能力。CcPAL4在褐化出现时(继代后17 d)上调表达,可能参与了致褐物质的合成。这种差异表达可能是由于不同的PAL基因对不同的信号和环境因素做出了响应,或者在不同的代谢途径中发挥了特定的作用[37]。
-
本研究表明:CcPAL蛋白具有酸性、稳定、亲水特性,可能参与基因转录调控等生物过程。系统进化分析显示:CcPAL5和CcPAL6与裸子植物关系更近。不同CcPAL基因在愈伤组织增殖中表现出不同表达模式,影响组织增殖和褐化。研究结果为理解山核桃生长发育和次生代谢物合成提供了重要信息,并为CcPAL在山核桃酚类物质合成、愈伤组织增殖与褐化、组织分化和器官再生中的作用机制研究奠定了基础,为山核桃遗传改良和产业发展提供了理论支持。
Genome-wide identification and expression profiling ofPALgene family inCarya cathayensis
-
摘要:
目的苯丙氨酸解氨酶(PAL)作为酚类物质合成代谢的起始酶,对植物的生长发育及环境适应起着至关重要的作用。鉴定山核桃 Carya cathayensis PAL( CcPALs)基因家族成员,分析家族成员在愈伤组织增殖褐化过程中的表达模式,为挖掘能够抑制褐化、促进再生的 PAL基因提供理论依据。 方法以山核桃全基因组数据为基础,通过比对筛选出包含PAL保守基序的PAL家族成员。运用生物信息学方法,对山核桃PAL蛋白的理化性质、基因结构等特征进行预测分析,并与多个物种的PALs序列共同构建发育进化树。同时,通过转录组测序,分析继代后愈伤组织中 CcPALs基因的表达变化,并制作表达热图。 结果山核桃共有6条PAL蛋白序列,长度为657~760个氨基酸。这些蛋白均属于酸性、稳定且亲水的蛋白,主要定位在细胞核和内质网上,蛋白二级结构以α-螺旋与无规则卷曲为主。进化关系分析表明:CcPAL1~CcPAL4聚类在双子叶植物分支,而CcPAL5和CcPAL6则聚类在裸子植物分支。表达分析显示: CcPALs基因在愈伤组织继代后呈现出不同的表达模式。 结论CcPALs成员之间具有较高的保守性,但CcPAL5和CcPAL6在进化关系上与裸子植物PAL更为接近,可能与CcPAL1~CcPAL4在次生代谢物合成过程中的功能存在较大差异。在愈伤组织增殖褐化过程中,进化树上不同分支的 CcPALs表达模式各异,可能控制着酚类物质的代谢流,进而影响山核桃的再生进程。图7表4参37 Abstract:ObjectivePhenylalanine ammonia-lyase (PAL), the initial enzyme in the biosynthesis of phenolic substances, is of crucial significance in plant growth, development, and environmental adaptation. The present study aims to identify the members of the PALgene family in Carya cathayensis( CcPALs) and analyze their expression patterns during callus proliferation and browning. This analysis may provide a theoretical basis for the manipulation of CcPALsto inhibit browning and promote regeneration. MethodBased on the whole-genome data of C. cathayensis, PAL family members containing conserved PAL motifs were screened. The physicochemical properties and gene-structure characteristics of CcPAL proteins were predicted and analyzed. A phylogenetic tree was constructed using PAL sequences from multiple species. Additionally, the expression changes of CcPALsin callus after subculture were examined through transcriptomic analysis. ResultThere were six PALs in C. cathayensisin total. The amino-acid lengths ranged from 657 to 760. These proteins were all acidic, stable, and hydrophilic, mainly localized in the nucleus and endoplasmic reticulum. The secondary structure of the protein was dominated by α-helix and random coil. Phylogenetic analysis revealed that CcPAL1−CcPAL4 cluster in the dicotyledonous plant branch, while CcPAL5 and CcPAL6 cluster in the gymnosperm branch. Expression analysis showed that the CcPALsdisplayed different expression patterns after callus subculture. ConclusionCcPALs exhibit high conservation. However, CcPAL5 and CcPAL6 were more closely related to gymnosperm PALs, potentially indicating significant functional differences from CcPAL1−CcPAL4 in secondary metabolism. During the callus proliferation and browning process, the expression patterns of CcPALsin different branches vary. This variation may potentially regulate the metabolic flux of phenolic substances and thus influence the regeneration process of C. cathayensis. [Ch, 7 fig. 4 tab. 37 ref.] -
图 2CcPAL家族的基因结构(A)、保守蛋白(B)、保守基序(C)和基序标识分析(D)
Figure 2Analysis of gene structure (A), conserved proteins (B), conserved motifs (C), and motif logos (D) of theCcPALfamily
图 6山核桃愈伤组织增殖过程中CcPALs基因表达热图
Figure 6Heat map ofCcPALsexpression during the proliferation of callus inC. cathayensis
图 7继代后不同时间下山核桃愈伤组织中的CcPALs基因表达
Figure 7Expression ofCcPALsgene in theC. cathayensiscallus at different time points after subculture
表 1实验中使用的RT-qPCR引物
Table 1.RT-qPCR primers used in the experiment
名称 上游引物(5′→3′) 下游引物(5′→3′) β-actin ATGGCCGATGCTGAGGATATT CACCAGTGTGTCGAGGTCTGC CcPAL1 ATGGGTTGCCTTCGAATCTC CCAGGGAGTTCACATCTTGG CcPAL2 AGGTTACTCGGGCATCAGAT GCCTATCAAAAGACCAGCGA CcPAL3 CTGCAGCGAAACCTATCCAT TGGCAGTGTTCAAGTTCCTC CcPAL4 CAGTGCGACATACCCATTGA TTCCTCAAAGACCCCGATCT CcPAL5 GCTTGGGGTAGAGGTAGCAA GCATTGGAGAAGTCACCGTG CcPAL6 TCAGGGGATCTTGTACCGTT GCAGAAATCCCAGCTCTTGT  下载:
导出CSV
下载:
导出CSV
表 2CcPALs蛋白理化性质
Table 2.Physicochemical properties of CcPALs protein
蛋白质 氨基酸数量/个 主要氨基酸组分(相对占比/%) 理论等电点 分子量/kDa 不稳定系数 脂肪指数 平均亲水性系数 CcPAL1 713 Leu(10.7)、Ala(9.8)、Gly(7.7) 6.41 77.54 32.96 91.85 −0.149 CcPAL2 657 Leu(11.4)、Ala(9.1)、Glu(7.3) 6.07 71.87 32.62 95.65 −0.114 CcPAL3 708 Leu(11.0)、Ala(9.2)、Gly(7.5) 6.23 77.34 32.40 92.75 −0.189 CcPAL4 710 Leu(10.3)、Ala(9.9)、Glu(8.0) 6.10 77.50 37.69 91.56 −0.160 CcPAL5 712 Leu(10.1)、Ala(8.6)、Glu(7.7) 6.01 78.57 34.77 90.27 −0.174 CcPAL6 760 Leu(10.3)、Ala(8.8)、Glu(7.5) 6.22 83.88 35.75 89.08 −0.214 说明:Leu. 亮氨酸; Ala. 丙氨酸; Gly. 甘氨酸; Glu. 谷氨酸。 下载:
导出CSV
表 3CcPALs蛋白亚细胞定位预测
Table 3.Subcellular localization prediction of CcPALs protein inC. cathayensis
蛋白质 预测定位 定位信号 膜类型 CcPAL1 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL2 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL3 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL4 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 CcPAL5 细胞核、内质网 核定位信号 外周、脂锚定 CcPAL6 细胞核、线粒体、内质网 核定位信号 外周、脂锚定 下载:
导出CSV
表 4CcPALs蛋白二级结构
Table 4.Secondary structure of CcPALs protein
蛋白质 占比/% α-螺旋 延伸链结构 β-转角 无规则卷曲 CcPAL1 53.02 8.70 6.73 31.56 CcPAL2 55.71 8.37 6.09 29.83 CcPAL3 55.37 7.49 6.50 30.65 CcPAL4 56.48 7.46 5.49 30.56 CcPAL5 55.34 9.13 6.88 28.65 CcPAL6 51.97 8.29 6.45 33.29 下载:
导出CSV
-
[1] SAITO K, YONEKURA-SAKAKIBARA K, NAKABAYASHI R,et al. The flavonoid biosynthetic pathway inArabidopsis: structural and genetic diversity [J].Plant Physiology and Biochemistry, 2013,72: 21−34. [2] HUANG Junli, GU Min, LAI Zhibing,et al. Functional analysis of theArabidopsisPALgene family in plant growth, development, and response to environmental stress [J].Plant Physiology, 2010,153(4): 1526−1538. [3] REICHERT A I, HE Xianzhi, DIXON R A. Phenylalanine ammonia-lyase (PAL) from tobacco (Nicotiana tabacum): characterization of the four tobaccoPALgenes and active heterotetrameric enzymes [J].Biochemical Journal, 2009,424(2): 233−242. [4] 曾嘉丽, 欧阳林娟, 刘家林, 等. 水稻PAL基因的全基因组分析及胁迫表达研究[J]. 基因组学与应用生物学, 2018,37(9): 3881−3888.ZENG Jiali, OUYANG Linjuan, LIU Jialin,et al. Whole genome analysis and stress expression research ofPALgene in rice [J].Genomics and Applied Biology, 2018,37(9): 3881−3888. [5] YAN Feng, LI Huaizhu, ZHAO Peng. Genome-wide identification and transcriptional expression of thePALgene family in common walnut (Juglans regiaL. )[J/OL].Genes, 2019,10(1): 46[2024-10-01]. DOI: 10.3390/genes10010046. [6] ROHDE A, MORREEL K, RALPH J,et al. Molecular phenotyping of the pal1 and pal2 mutants ofArabidopsis thalianareveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism [J].The Plant Cell, 2004,16(10): 2749−2771. [7] LI Guohui, SONG Cheng, MANZOOR M A,et al. Functional and kinetics of two efficient phenylalanine ammonia lyase fromPyrus bretschneideri[J/OL].BMC Plant Biology, 2023,23(1): 612[2024-10-01]. DOI: 10.1186/s12870-023-04586-0. [8] BAGAL U R, LEEBENS-MACK J H, LORENZ W W,et al. The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage[J/OL].BMC Genomics, 2012,13(suppl 3): S1[2024-10-01]. DOI: 10.1186/1471-2164-13-S3-S1235. [9] PANT S, HUANG Yinghua. Genome-wide studies ofPALgenes in sorghum and their responses to aphid infestation[J/OL].Scientific Reports, 2022,12(1): 22537[2024-10-01]. DOI: 10.1038/s41598-022-25214-1. [10] 谢林峰, 凌晓晓, 黄圣妍, 等. 临安区山核桃林地土壤水解酶活性空间分布特征及土壤肥力评价[J]. 浙江农林大学学报, 2022,39(3): 625−634.XIE Linfeng, LING Xiaoxiao, HUANG Shengyan,et al. Spatial distribution characteristics of soil hydrolase activities and soil fertility evaluation ofCarya cathayensisforests in Lin’an District [J].Journal of Zhejiang A&F University, 2022,39(3): 625−634. [11] 张璐璐, 贾桂民, 叶建丰, 等. 浙江临安山核桃干腐病发生发展规律[J]. 浙江农林大学学报, 2013,30(1): 148−152.ZHANG Lulu, JIA Guimin, YE Jianfeng,et al. Frequency ofCarya cathayensiscanker disease in Lin’an City, Zhejiang Province [J].Journal of Zhejiang A&F University, 2013,30(1): 148−152. [12] LIU Chen, FAN Hongrui, ZHANG Jiaqi,et al. Combating browning: mechanisms and management strategies inin vitroculture of economic woody plants[J/OL].Forestry Research, 2024,4: e032[2024-10-01]. DOI: 10.48130/forres-0024-0026. [13] NAGY E Z A, TORK S D, LANG P A,et al. Mapping the hydrophobic substrate binding site of phenylalanine ammonia-lyase fromPetroselinum crispum[J].ACS Catalysis, 2019,9(9): 8825−8834. [14] TOMOIAGĂ R B, TORK S D, FILIP A,et al. Phenylalanine ammonia-lyases: combining protein engineering and natural diversity [J].Applied Microbiology and Biotechnology, 2023,107(4): 1243−1256. [15] XU Feng, DENG Guang, CHENG Shuiyuan,et al. Molecular cloning, characterization and expression of the phenylalanine ammonia-lyase gene fromJuglans regia[J].Molecules, 2012,17(7): 7810−7823. [16] CHEN Chengjie, CHEN Hao, ZHANG Yi,et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J].Molecular Plant, 2020,13(8): 1194−1202. [17] HUANG Youjun, XIAO Lihong, ZHANG Zhongren,et al. The genomes of pecan and Chinese hickory provide insights intoCaryaevolution and nut nutrition[J/OL].GigaScience, 2019,8(5): giz036[2024-10-01]. DOI: 10.1093/gigascience/giz036. [18] PAYSAN-LAFOSSE T, BLUM M, CHUGURANSKY S,et al. InterPro in 2022 [J].Nucleic Acids Research, 2023,51(D1): D418−D427. [19] GASTEIGER E, HOOGLAND C, GATTIKER A,et al. Protein identification and analysis tools on the ExPASy server[M]// WALKER J M.The Proteomics Protocols Handbook. Totowa N J: Humana Press, 2005: 571−607. [20] THUMULURI V, ARMENTEROS J J A, JOHANSEN A R,et al. DeepLoc 2.0: multi-label subcellular localization prediction using protein language models [J].Nucleic Acids Research, 2022,50(W1): W228−W234. [21] GEOURJON C, DELÉAGE G. SOPMA: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments [J].Computer Applications in the Biosciences, 1995,11(6): 681−684. [22] KELLEY L A, MEZULIS S, YATES C M,et al. The Phyre2 web portal for protein modeling, prediction and analysis [J].Nature Protocols, 2015,10(6): 845−858. [23] BAILEY T L, JOHNSON J, GRANT C E,et al. The MEME suite [J].Nucleic Acids Research, 2015,43(W1): W39−W49. [24] TAMURA K, STECHER G, KUMAR S. MEGA11: molecular evolutionary genetics analysis version 11 [J].Molecular Biology and Evolution, 2021,38(7): 3022−3027. [25] OKADA T, MIKAGE M, SEKITA S. Molecular characterization of the phenylalanine ammonia-lyase fromEphedra sinica[J].Biological & Pharmaceutical Bulletin, 2008,31(12): 2194−2199. [26] WANNER L A, LI Guoqing, WARE D,et al. The phenylalanine ammonia-lyase gene family inArabidopsis thaliana[J].Plant Molecular Biology, 1995,27(2): 327−338. [27] 施圆圆, 张声祥, 赵德蕊, 等. 异叶天南星苯丙氨酸解氨酶基因的克隆与蛋白结构分析[J]. 植物科学学报, 2019,37(2): 221−229.SHI Yuanyuan, ZHANG Shengxiang, ZHAO Derui,et al. Gene cloning and structure characterization of phenylalanine ammonia-lyase fromArisaema heterophyllum[J].Plant Science Journal, 2019,37(2): 221−229. [28] LIN Weiping, LIU Ao, WENG Caihong,et al. Cloning and characterization of a novel phenylalanine ammonia-lyase gene fromInonotus baumii[J].Enzyme and Microbial Technology, 2018,112: 52−58. [29] DONG Chunjuan, CAO Ning, ZHANG Zhigang,et al. Phenylalanine ammonia-lyase gene families in cucurbit species: structure, evolution, and expression [J].Journal of Integrative Agriculture, 2016,15(6): 1239−1255. [30] SHANG Qingmao, LI Liang, DONG Chunjuan. Multiple tandem duplication of the phenylalanine ammonia-lyase genes inCucumis sativusL. [J].Planta, 2012,236(4): 1093−1105. [31] DONG Chunjuan, SHANG Qingmao. Genome-wide characterization of phenylalanine ammonia-lyase gene family in watermelon (Citrullus lanatus) [J].Planta, 2013,238(1): 35−49. [32] YANG Ji, HUANG Jinxia, GU Hongya,et al. Duplication and adaptive evolution of theChalconesynthase genes ofDendranthema(Asteraceae) [J].Molecular Biology and Evolution, 2002,19(10): 1752−1759. [33] WU Zhihua, GUI Songtao, WANG Shuzhen,et al. Molecular evolution and functional characterisation of an ancient phenylalanine ammonia-lyase gene (NnPAL1) fromNelumbo nucifera: novel insight into the evolution of the PAL family in angiosperms[J/OL].BMC Evolutionary Biology, 2014,14: 100[2024-10-01]. DOI: 10.1186/1471-2148-14-100. [34] HAMBERGER B, ELLIS M, FRIEDMANN M,et al. Genome-wide analyses of phenylpropanoid-related genes inPopulus trichocarpa, Arabidopsis thaliana, andOryza sativa: thePopuluslignin toolbox and conservation and diversification of angiosperm gene families [J].Canadian Journal of Botany, 2007,85(12): 1182−1201. [35] LI Guohui, WANG Han, CHENG Xi,et al. Comparative genomic analysis of thePALgenes in five Rosaceae species and functional identification of Chinese white pear[J/OL].PeerJ, 2019,7: e8064[2024-10-01]. DOI: 10.7717/peerj.8064. [36] AMJAD M, WANG Yuexia, HAN Shiming,et al. Genome wide identification of phenylalanine ammonia-lyase (PAL) gene family inCucumis sativus(cucumber) against abiotic stress[J/OL].BMC Genomic Data, 2024,25(1): 76[2024-10-01]. DOI 10.1186/s12863-024-01259-1. [37] RASOOL F, UZAIR M, NAEEM M K,et al. Phenylalanine ammonia-lyase (PAL) genes family in wheat (Triticum aestivumL. ): genome-wide characterization and expression profiling[J/OL].Agronomy, 2021,11(12): 2511[2024-10-01]. DOI: 10.3390/agronomy11122511. -

- 链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240587
 点击查看大图
点击查看大图

计量
- 文章访问数:712
- HTML全文浏览量:216
- PDF下载量:104
- 被引次数:0
